Research
Network mechanisms of cognitive control: We study how brain networks interact to transmit, select, or inhibit information for flexible human behaviors.
Integration of control representations: We investigate the structure, dynamics, and interactions between task representations that facilitate cognitive control.
Methods: Our research utilizes a variety of methods:
Cognitive paradigms and computational modeling (e.g., Bayesian generative model) to constrain our neuroimaging analyses.
Multimodal neuroimaging, fMRI, EEG, time-frequency and connectivity analyses.
Representational analyses, decoding and represention similarity analyses.
Experimental neuropsychology, lesion approaches, and causal TMS manipulations.
Structure and dynamics of task representations
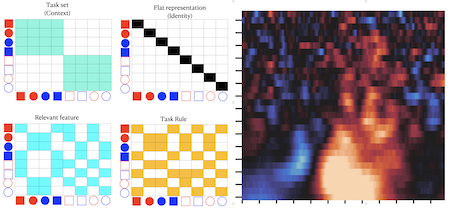
Task representation is a central component of cognitive control. However, several important properties of task representations are not well understood, including their structure and interactive dynamics. This is likely because it is difficult to directly observe these latent properties of mental representations using conventional behavioral and neuroimaging methods. We are conducting mulitmodal neuroimag, lesion, and behavioral modeling studies to discover the structure and dynamics of representations that facilitate cognitive control.
Mechanisms of cognitive integration
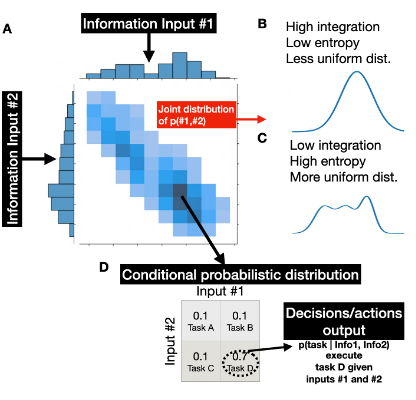
Many cognitively challenging tasks require the integration of diverse information sources rather than relying on a single factor. For example, consider a chef preparing a complex dish in a restaurant kitchen. The chef must integrate the recipe (long-term memory), the ticket order (working memory), and the progress of the dish on the stove (sensory input). Thus, behavior is continuously shaped by the integration of multiple factors. Despite its clear behavioral significance, the term “integration” is frequently employed in the field loosely without specificity regarding the underlying cognitive and neural mechanisms. We are currently developing a neurocognitive model of cognitive integration.
Integrative functions of working memory
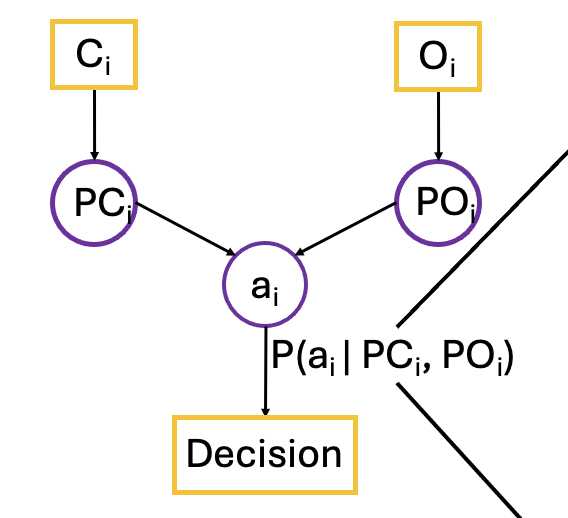
Many have argued that working memory is the medium that guides cognitive control. However, beyond estimating the precision and capacity of working memory content, it is unclear how humans actively and adaptively use working memory to guide our actions and decisions. Our lab is actively developing computational models to determine how working memory integrates diverse sources of information for decision-making. This work will integrate modeling, multimodal neuroimaging, decoding, model-based fMRI/EEG analyses, and linear state-space modeling.
Cognitive flexibility and hierarchical cognitive control
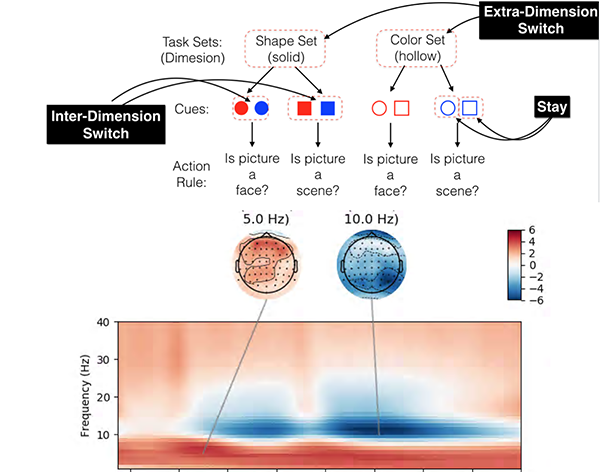
The cardinal characteristic of cognitive control is it is flexible and context dependent. We are not governed by the same set of rules in every situation. If so, how do we use circumstantial information to adjust our actions, specifically, adjust mappings between sensory and motor processes? We have developed a paradigm that requires human subjects to switch between action rules depending on a superordinate, circumstantial context. Parallel EEG and fMRI studies are now in progress.
Spectral neural dynamics and cognitive control
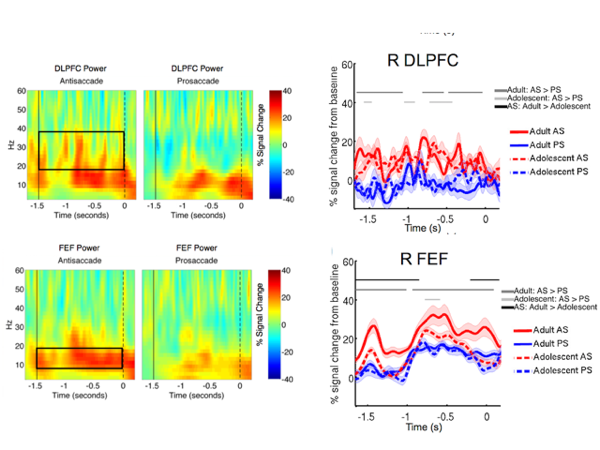
How do brain networks flexibly process and communicate information? Possibly through oscillatory neural activities. Different brain rhythms are thought to reflect distinct biophysical and circuit-level processes, thus could be indices of distinct neurocognitive mechanisms. We study how oscillatory neural dynamics support cognitive control in adults and during development.
Cognitive control functions of the human thalamocortical system
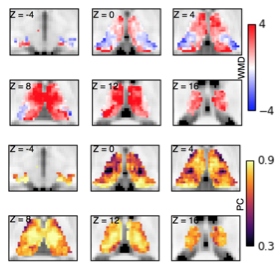
The mammalian brain can be conceptualized as a thalamocortical system, yet the thalamus is often ignored in studies of brain network organization. One major focus of our research is to use advanced network neurosciecne techniques to investigate the thalamus's contribution to diverse network and cognitive functions.
Effects of thalamic lesions
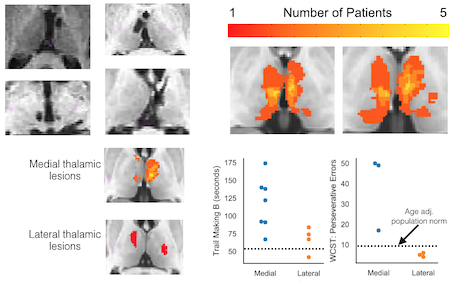
The human thalamus is a difficult region to study because of its small size and deep location. To address this challenge, we are now conducting studies with a novel approach: combining multimodal neuroimaging (fMRI and EEG) and human lesions studies. The goal is to determine how the disruption of thalamocortical interactions after thalamic lesion affects cortical neural activity and behavior. We are now focusing on how the thalamus modulate cortical evoked responses, neural oscillations, and cortico-cortical functional connectivity for cognitive control.
Netowrk processes implementing cognitive control
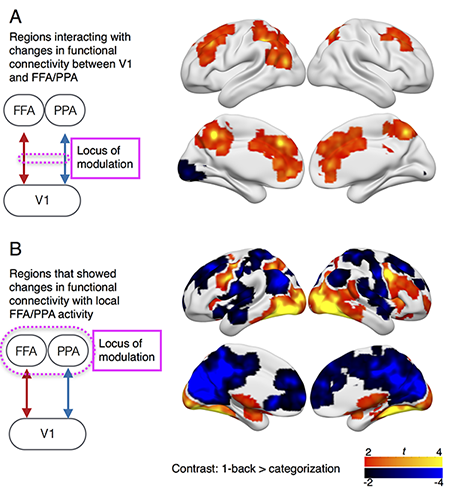
Could functional connectivity be modulated by "top-down biasing signals"? We are studying how cognitive control influences information exchange between task-related brain regions, and to identify brain regions interacting with dynamic functional connectivity patterns. We use TMS to causally map regions that provide "biasing signals" to enhance or inhibit functional connectivity for cognitive control.